

Origin and content of alkenone of the Early Miocene marine sediments from IODP U1501 in the South China Sea
-
摘要:
海洋沉积物中的长链烯酮由海洋单细胞钙化藻类颗石藻生产,是一种被广泛应用于古气候研究领域中的分子标记物。长链烯酮碳同位素是重建地质历史时期海水、大气CO2浓度的可靠方法之一。在此方法中,需要利用颗石大小对颗石藻生理参数b值进行修正,因此需要厘清哪类颗石藻对烯酮的贡献是一个重要的科学问题。目前认为新生代海洋沉积物中主要的长链烯酮生产者为Noelaerhabdaceae科的颗石藻,包含Emiliania huxleyi, Gephyrocapsa spp., Reticulofenestra spp., Cyclicargolithus spp.,但对它们具体的贡献程度仍然未知。因此,本文以南海国际大洋发现计划IODP U1501站早中新世海洋沉积物为研究材料,对比了沉积物中颗石与烯酮的绝对含量,发现Cyclicargolithus属的颗石丰度与烯酮含量具有显著的相关性(r=0.44,p<0.01),而Reticulofenestra spp.的相关性较弱(r=0.09,p=0.5)。研究认为早中新世长链烯酮的主要生产者为Cyclicargolithus属,Reticulofenestra属次之。此外,研究还测量了各颗石属种的长度,并估算了颗石藻细胞的直径。研究认为Cyclicargolithus属对烯酮的高贡献源于其较大的细胞直径。从长链烯酮生产者的种类、细胞体积及其生命过程处着眼,有利于评估应用碳同位素分馏重建海洋中CO2浓度的可行性,对准确地还原地质历史时期大气CO2浓度具有重要作用。
-
关键词:
- 长链烯酮 /
- 颗石藻 /
- 早中新世 /
- IODP U1501站 /
- 南海
Abstract:Alkenones are ubiquitous in ocean sediments and have been widely used in paleoceanography and paleoclimatology. Alkenones can be used to reconstruct the CO2 concentration of seawater in geological history, which is key to understanding the mechanism of climate evolution and predicting future climate change. In modern open ocean environments, the main producers of alkenones are Noelaerhabdaceae, including the calcifying haptophytes (coccolithophores) Emiliania huxleyi and Gephyrocapsa spp. It is generally believed that the most reliable producers of alkenones in the Cenozoic marine sediments belong to genus Reticulofenestra of Family Noelaerhabdaceae. By comparing the absolute abundance of coccoliths and alkenones in the Early Miocene sediments at IODP Site U1501 in the South China Sea, it is found that the abundance of Cyclicargolithus spp. coccoliths and alkenones are well correlated (r=0.44, p<0.01). Therefore, we believe that Cyclicargolithus spp. is the main producer of alkenones in the Early Miocene, followed by Reticulofenestra spp. In addition, we measured the coccolith length of each species and estimate the diameter of coccolithophores cells. We inferred that the high contribution of Cyclicargolithus spp. to alkenones is due to its larger cell diameter. Our finding is helpful to evaluate the feasibility of using carbon isotope fractionation to reconstruct the atmospheric CO2 concentration in geological history.
-
Key words:
- alkenones /
- coccolithophore /
- Early Miocene /
- IODP U1501 /
- South China Sea
-
-
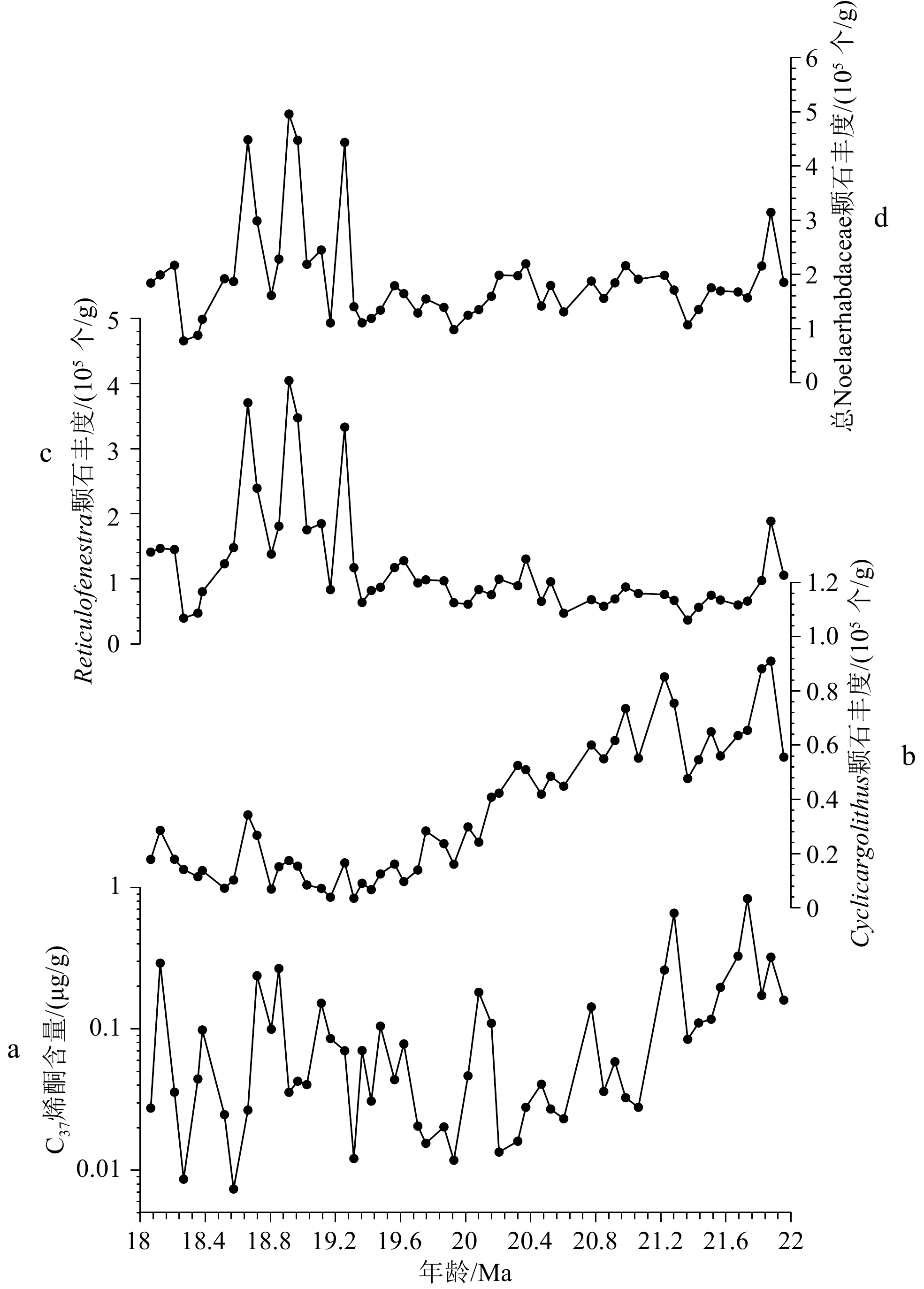
图 3 南海IODP站位U1501C早中新世18~22 Ma的C37长链烯酮含量(a),Cyclicargolithus属颗石丰度(b),Reticulofenestra属颗石丰度(c)以及总Noelaerhabdaceae颗石丰度(d)
Figure 3.
表 1 颗石绝对丰度与烯酮浓度之间Pearson相关性分析结果
Table 1. Pearson correlation analysis between the coccolith abundance and the alkenone concentration
相关系数r p值 Noelaerhabdaceae科颗石的总丰度 0.005 315 0.969 87 Cyclicargolithus属颗石丰度 0.443 09 0.000 892 Reticulofenestra颗石丰度 0.093 14 0.507 13  下载: 导出CSV
下载: 导出CSV
-
[1] Marlowe I T, Brassell S C, Eglinton G, et al. , Long chain unsaturated ketones and esters in living algae and marine sediments [J], Organic Geochemistry, 1984, 6(1): 135-141.
[2] Volkman J K, Barrerr S M, Blackburn S I, et al. , Alkenones in Gephyrocapsa oceanica: Implications for studies of paleoclimate [J], Geochimica Et Cosmochimica Acta, 1995, 59(3): 513-520.
[3] Volkman J K, Eglnton G, Corner E D S, et al. Long-chain alkenes and alkenones in the marine coccolithophorid Emiliania huxleyi [J]. Phytochemistr, 1980, 19(12): 2619-2622. doi: 10.1016/S0031-9422(00)83930-8
[4] Marlowe I T, Brassell S C, Eglinton G, et al. , Long-chain alkenones and alkyl alkenoates and the fossil coccolith record of marine sediments [J], Chemical Geology, 1990, 88(3): 349-375.
[5] Brassell S C, Eglinton G, Marlowe I T, et al. , Molecular stratigraphy: a new tool for climatic assessment [J], Nature, 1986, 320(6058): 129-133.
[6] Jasper J and Hayes J, A carbon isotope record of CO2 level during the late Quaternary [J], Nature, 1990, 347(6292): 462-4.
[7] Eglinton G, Stuart B, Antoni R, et al. , Molecular record of secular sea surface temperature changes on 100-year timescales for glacial terminations I, II and IV [J], Nature, 1992, 356(6368): 423-426.
[8] Bard E, Frauke R and Corinne S, Interhemispheric synchrony of the last deglaciation inferred from alkenone palaeothermometry [J], Nature, 1997, 385(6618): 707-710.
[9] Cacho I, Joan O G, Carles P, et al. , Dansgaard-Oeschger and Heinrich event imprints in Alboran Sea paleotemperatures [J], Paleoceanography, 1999, 14(6): 698-705.
[10] Bolton C, Kira L, Samantha G, et al. , Glacial–interglacial productivity changes recorded by alkenones and microfossils in late Pliocene eastern equatorial Pacific and Atlantic upwelling zones [J], Earth and Planetary Science Letters, 2010, 295(3-4): 401-411.
[11] Prahl F G and Wakeham S G. Calibration of unsaturation patterns in long-chain ketone compositions for palaeotemperature assessment [J], Nature, 1987, 330(6146): 367-369.
[12] Müller P J. , Kirst G, Ruhland G, et al. , Calibration of the alkenone paleotemperature index UK37′ based on core-tops from the eastern South Atlantic and the global ocean (60°N-60°S) [J], Geochimica et Cosmochimica Acta, 1998, 62(10): 1757-1772.
[13] Eltgroth M L, Watwood R L and Wolfe G V. Production and cellular localization of neutral long-chain lipids in the Haptophyte algae Isochrysis galbana and Emiliania huxleyi [J], Journal of Phycology, 2005, 41(5): 1000-1009.
[14] Pahnke K and Sachs J P. Sea surface temperatures of southern midlatitudes 0–160 kyr B. P [J], Paleoceanography, 2006, 21(2): 1-17.
[15] Jasper J, Hayes J, Mix A, et al. , Photosynthetic fractionation of 13C and concentrations of dissolved CO2 in the central equatorial Pacific during the last 255 000 years [J], Paleoceanography, 1994, 9(6): 781-98.
[16] Bidigare R R, Fluegge A, Freeman K H, et al. , Consistent fractionation of 13C in nature and in the laboratory: Growth-rate effects in some haptophyte algae [J], Global Biogeochemical Cycles, 1997, 11(2): 279-292.
[17] Pagani M, Arthur M A and Freeman K H. Miocene evolution of atmospheric carbon dioxide [J], Paleoceanography, 1999, 14(3): 273-292.
[18] Pagani M. The alkenone-CO2 proxy and ancient atmospheric carbon dioxide [J], Philosophical transactions. Series A, Mathematical, physical, and engineering sciences, 2002, 360(1793): 609-632.
[19] Henderiks J and Mark P. Refining ancient carbon dioxide estimates: Significance of coccolithophore cell size for alkenone-based pCO2 records [J], Paleoceanography, 2007, 22(3): 324-329.
[20] Seki O, Foster G L, Schmidt D N, et al. , Alkenone and boron-based Pliocene pCO2 records [J], Earth and Planetary Science Letters, 2010, 292(1-2): 201-211.
[21] Farrimond P, Eglinton G and Brassell S C. Alkenones in Cretaceous black shales, Blake-Bahama Basin, western North Atlantic [J], Organic Geochemistry, 1986, 10(4-6): 897-903.
[22] Brassell S C and Mirela D. Recognition of alkenones in a lower Aptian porcellanite from the west-central Pacific [J], Organic Geochemistry, 2004, 35(2): 181-188.
[23] Plancq J, Grossi V, Henderiks J, et al. , Alkenone producers during late Oligocene–early Miocene revisited [J], Paleoceanography, 2012, 27(1): PA1202.
[24] Luo Y. et al., Invariance of the carbonate chemistry of the South China Sea from the glacial period to the Holocene and its implications to the Pacific Ocean carbonate system [J]. Earth Planet. Sci. Lett., 2018, 492(1): 112-120.
[25] Jian Z M, Jin H Y, Kaminski M A, et al. , Discovery of the marine Eocene in the northern South China Sea [J], National Science Review, 2019, 6(5): 881-885.
[26] Henderiks J and Mark P. Coccolithophore cell size and the Paleogene decline in atmospheric CO2 [J], Earth and Planetary Science Letters, 2008, 269(3-4): 576-584.
[27] Young J. Size variation of Neogene Reticulofenestra coccoliths from Indian Ocean DSDP Cores [J], Journal of Micropalaeontology, 1990, 9(1): 71-85.
[28] Aubry M. Paleogene calcareous nannofossil stratigraphy of ODP Leg 120 sites [J], PANGAEA , 1992, 120(1): 471-491.
[29] Beaufort L L. Size Variations in Late Miocene Reticulofenestra and implication for paleo climatic interpretation [J], Memorie di Scienze Geologiche, 1992, 43(1): 339-350.
[30] Tappan H N. The Paleobiology of Plant Protists [J]. Geologiska Fö reningeni Stockholm Fö rhandlingar, 1980, 104(2): 156.
[31] Perch-Nielsen K. Morphological Description of Calcareous Nannofossils Assemblage of a Middle-Miocene to Late-Miocene Section in the Niger-Delta, Nigeria [J]. Open Journal of Geology, 1985, 8(9): 427-554.
[32] Young, J. R. , Bown P. R. , Lees J. A. , 2022, Nannotax3 website. International Nannoplankton Association. Accessed 21 Apr. 2022. URL: www. mikrotax. org/Nannotax3
[33] Bordiga M, Bartol M and Henderiks J. Absolute nannofossil abundance estimates: Quantifying the pros and cons of different techniques [J], Revue de Micropaléontologie, 2015, 58(3): 155-165.
[34] Šupraha L and Henderiks J. A 15-million-year-long record of phenotypic evolution in the heavily calcified coccolithophore Helicosphaera and its biogeochemical implications [J], Biogeosciences, 2020, 17(11): 2955-2969.
[35] Prahl F G, Wakeham S G. Calibration of unsaturation patterns in long-chain ketone compositions for paleo temperature assessment [J]. Nature, 1987, 330(6146): 367-369. doi: 10.1038/330367a0
[36] Brassell S C, Climatic influences on the Paleogene evolution of alkenones [J], Paleoceanography, 2014, 29(3): 255-272.
[37] Barnes P J, Brassell S C, Comet P, et al. , Preliminary lipid analyses of Core Sections 18, 24 and 30 from Hole 402A [J]. Init. Rep. Deep Sea Drill. Proj, 1979, 48(1): 965-976.
[38] Nicole R. Marshall, Anne de Vernal, Alfonso Mucci, Markus Kienast, Alexandra Filippova, Claude Hillaire-Marcel, Carbonate dissolution and environmental parameters govern coccolith vs. alkenone abundances in surface sediments from the northwest North Atlantic [J], Marine Micropaleontology, 2021, 169(4): 102032
[39] 王乙晶, 金海燕, 翦知湣, 徐娟. 南海北部晚渐新世与早中新世之交T60构造运动的古水深响应[J]. 海洋学报, 2021, 43(5):79-87
Wang yijing, Jin Haiyan, Jian Zhimin, et al. The response of paleo-water depth to T60 tectonic movement in the northern South China Sea during the late Oligocene to early Miocene [J]. Haiyang Xuebao, 2021, 43(5): 79-87.
[40] Zachos, J. , Pagani, M., Sloan, L., Thomas, E., and Billups, K. Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present [J]. Science, 2001, 292(5517): 686-693. doi: 10.1126/science.1059412
[41] Kameo K and Toshiaki T, Biostratigraphic significance of sequential size variations of the calcareous nannofossil genus Reticulofenestra in the Upper Pliocene of the North Atlantic [J], Marine Micropaleontology, 1999, 37(1): 41-52.
[42] Backman J and Hermelin J O R, Morphometry of the Eocene nannofossil Reticulofenestra umbilicus lineage and its biochronological consequences [J], Palaeogeography, Palaeoclimatology, Palaeoecology, 1986, 57(1): 103-116.
[43] Laws E A. , Popp B N, Bidigare R R, et al. , Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2]aq: Theoretical considerations and experimental results [J], Geochimica et Cosmochimica Acta, 1995, 59(6): 1131-1138.
[44] Popp B N, Laws E A, Bidigare R R, et al. , Effect of Phytoplankton Cell Geometry on Carbon Isotopic Fractionation [J], Geochimica et Cosmochimica Acta, 1998, 62(1): 69-77.
-


图(4)
表(1)
计量
- 文章访问数: 1244
- PDF下载数: 178
- 施引文献: 0