

Comparison in Sr/Ca ratios of different morphotypes of surface dwelling planktonic foraminifera species
-
摘要:
浮游有孔虫表层水种Globigerinoides ruber (白色)和Trilobatus sacculifer壳体是过去海洋环境信息的重要载体,其壳体Sr/Ca比值在重建古海洋环境变化中具有较大潜力,因此本文旨在探讨G. ruber(白色)和T. sacculifer的不同形态类型Sr/Ca比的差异及其影响因素。对取自西菲律宾海的MD06-3047B沉积物岩芯中G. ruber的两种形态类型,即G. ruber(sensu stricto, s.s.)和G. ruber(sensu lato, s.l.),以及T. sacculifer 的两种形态类型,即T. sacculifer (with sac)和T. sacculifer (without sac)壳体的Sr/Ca进行了测试分析。结果显示,MIS 3期(~48 ka)以来,MD06-3047B岩芯中G. ruber s.s.和G. ruber s.l. 的Sr/Ca差异较小;而T. sacculifer (with sac)和T. sacculifer (without sac)的Sr/Ca存在显著差异,且前者显著高于后者。通过与同站位表层海水温度和盐度等记录的对比,发现研究区G.ruber s.s.、G. ruber s.l.和T. sacculifer (without sac)壳体的Sr/Ca可能主要受控于海水温度,而T. sacculifer (with sac)壳体Sr/Ca受到海水盐度的影响。
-
关键词:
- 浮游有孔虫 /
- 形态类型 /
- G. ruber (white) /
- T. sacculifer /
- Sr/Ca
Abstract:Surface-dwelling foraminifera species Globigerinoides ruber (white) and Trilobatus sacculifer are the main proxy carriers for past oceanic environments. The Sr/Ca ratios of foraminifera play an important role in reflecting paleo-oceanic environmental changes. To understand the potential differences between the Sr/Ca ratios of these different morphotypes of G. ruber (white) and T. sacculifer and relevant affecting factors, we compared the Sr/Ca measurements of the two basic morphotypes of G. ruber (G. ruber sensu stricto and G. ruber sensu lato) and T. sacculifer (with sac and without sac) via the analysis of surface and downcore samples of sediment from Core MD06-3047B in the Western Philippine Sea. Results reveal that the Sr/Ca ratios of the two morphotypes of G. ruber (white) show a little difference since MIS 3 (~48 ka), while those of two types of T. sacculifer are significantly different from each other, of which that of T. sacculifer (without sac) is generally greater than that of the with-sac type. By comparing the seawater temperature and salinity from the same site, we found that the Sr/Ca ratios of G. ruber s.s., G. ruber s.l. and T. sacculifer (without sac) in our study area might be affected mainly by seawater temperature, and those of T. sacculifer (with sac) are mainly affected by surface salinity.
-
Key words:
- planktonic foraminifera /
- morphotype /
- G. ruber (white) /
- T. sacculifer /
- Sr/Ca
-
-
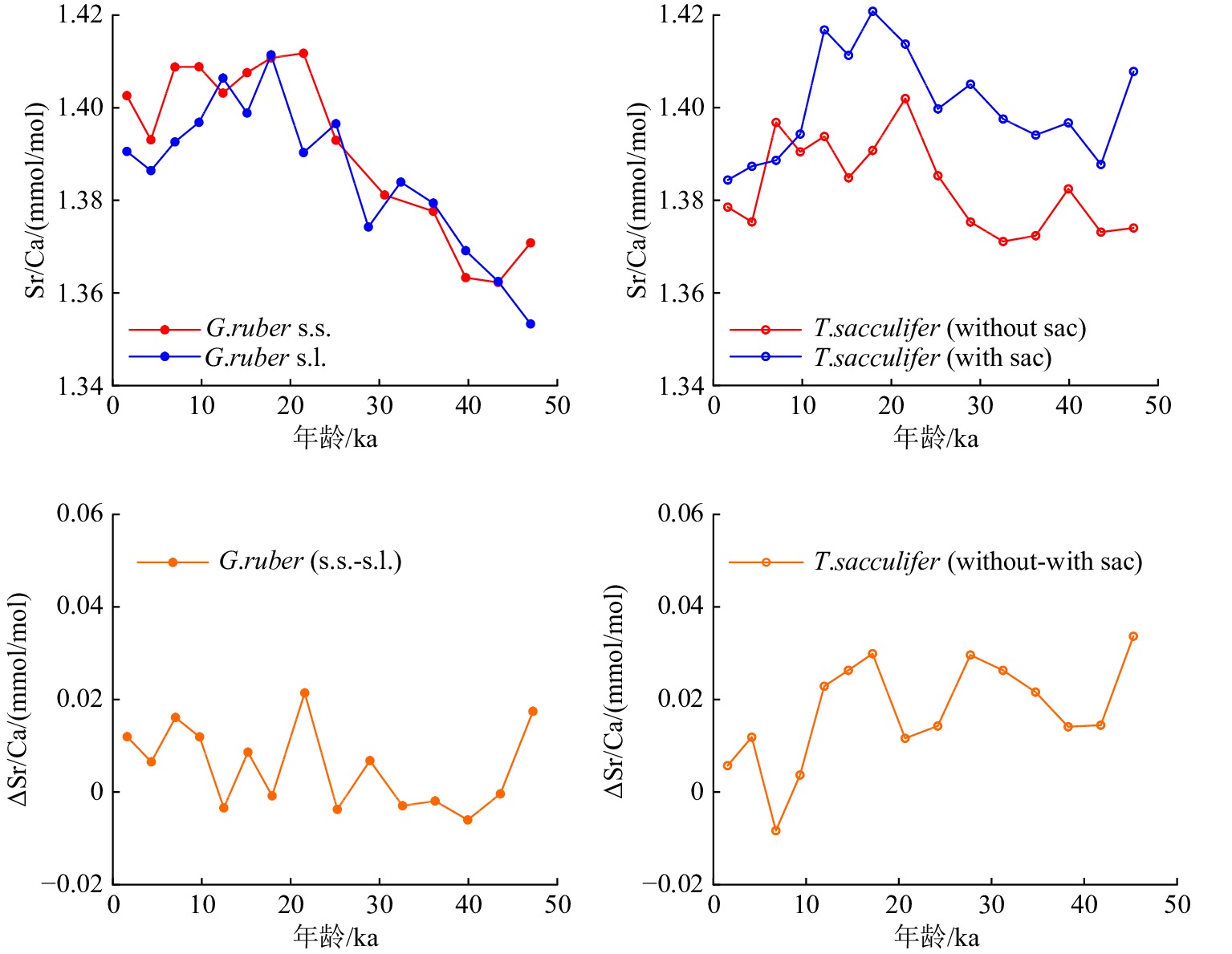
图 3 MD06-3047B孔G. ruber s.s.、G. ruber s.l.、T. sacculifer(with sac)和T. sacculifer(without sac)壳体的Sr/Ca比值,以及G. ruber和T. sacculifer不同形态类型壳体Sr/Ca差值
Figure 3.
表 1 MD06-3047B孔G. ruber s.s.和G. ruber s.l.以及T. sacculifer(with sac)和T. sacculifer(without sac)壳体Sr/Ca平均值(mmol/mol)以及韦尔奇检验结果(p<0.05)
Table 1. Mean shell Sr/Ca (mmol/mol) of G. ruber s.s., G. ruber s.l., T. sacculifer (with sac), and T. sacculifer (without sac) of Core MD06-3047B with the results of the Welch’s t-test at p<0.05 level
Sr/Ca Sr/Ca G.ruber s.s. 1.393 G.sacculifer (without sac) 1.383 G.ruber s.l. 1.387 G.sacculifer (with sac) 1.401 H H0 H Ha 注:其中H = H0表示接受虚假设,H = Ha表示拒绝虚假设。  下载: 导出CSV
下载: 导出CSV
-
[1] Mohtadi M, Steinke S, Lückge A, et al. Glacial to Holocene surface hydrography of the tropical eastern Indian Ocean[J]. Earth and Planetary Science Letters, 2010, 292(1-2): 89-97. doi: 10.1016/j.jpgl.2010.01.024
[2] Stott L, Poulsen C, Lund S, et al. Super ENSO and global climate oscillations at millennial time scales[J]. Science, 2002, 297(5579): 222-226. doi: 10.1126/science.1071627
[3] Koutavas A, Joanides S. El Niño-southern oscillation extrema in the Holocene and Last Glacial Maximum[J]. Paleoceanography, 2012, 27(4): PA4208.
[4] Steinke S, Chiu H Y, Yu P S, et al. Mg/Ca ratios of two Globigerinoides ruber (white) morphotypes: Implications for reconstructing past tropical/subtropical surface water conditions[J]. Geochemistry, Geophysics, Geosystems, 2005, 6(11): Q11005.
[5] Gussone N, Eisenhauer A, Tiedemann R, et al. Reconstruction of Caribbean Sea surface temperature and salinity fluctuations in response to the Pliocene closure of the Central American Gateway and radiative forcing, using δ44/40Ca, δ18O and Mg/Ca ratios[J]. Earth and Planetary Science Letters, 2004, 227(3-4): 201-214. doi: 10.1016/j.jpgl.2004.09.004
[6] Wang L J. Isotopic signals in two morphotypes of Globigerinoides ruber (white) from the South China Sea: implications for monsoon climate change during the last glacial cycle[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 161(3-4): 381-394. doi: 10.1016/S0031-0182(00)00094-8
[7] Kawahata H. Stable isotopic composition of two morphotypes of Globigerinoides ruber (white) in the subtropical gyre in the North Pacific[J]. Paleontological Research, 2005, 9(1): 27-35. doi: 10.2517/prpsj.9.27
[8] Antonarakou A, Kontakiotis G, Mortyn P G, et al. Biotic and geochemical (δ18O, δ13C, Mg/Ca, Ba/Ca) responses of Globigerinoides ruber morphotypes to upper water column variations during the last deglaciation, Gulf of Mexico[J]. Geochimica et Cosmochimica Acta, 2015, 170: 69-93. doi: 10.1016/j.gca.2015.08.003
[9] Steinke S, Kienast M, Groeneveld J, et al. Proxy dependence of the temporal pattern of deglacial warming in the tropical South China Sea: toward resolving seasonality[J]. Quaternary Science Reviews, 2008, 27(7-8): 688-700. doi: 10.1016/j.quascirev.2007.12.003
[10] Wara M W, Ravelo A C, Delaney M L. Permanent El Niño-like conditions during the Pliocene warm period[J]. Science, 2005, 309(5735): 758-761. doi: 10.1126/science.1112596
[11] Elderfield H, Vautravers M, Cooper M. The relationship between shell size and Mg/Ca, Sr/Ca, δ18O, and δ13C of species of planktonic foraminifera[J]. Geochemistry, Geophysics, Geosystems, 2002, 3(8): 1-13.
[12] André A, Weiner A, Quillévéré F, et al. The cryptic and the apparent reversed: lack of genetic differentiation within the morphologically diverse plexus of the planktonic foraminifer Globigerinoides sacculifer[J]. Paleobiology, 2013, 39(1): 21-39. doi: 10.1666/0094-8373-39.1.21
[13] Anand P, Elderfield H, Conte M H. Calibration of Mg/Ca thermometry in planktonic foraminifera from a sediment trap time series[J]. Paleoceanography, 2003, 18(2): 1050.
[14] Stoll H M, Schrag D P, Clemens S C. Are seawater Sr/Ca variations preserved in Quaternary foraminifera?[J]. Geochimica et Cosmochimica Acta, 1999, 63(21): 3535-3547. doi: 10.1016/S0016-7037(99)00129-5
[15] Lea D W, Mashiotta T A, Spero H J. Controls on magnesium and strontium uptake in planktonic foraminifera determined by live culturing[J]. Geochimica et Cosmochimica Acta, 1999, 63(16): 2369-2379. doi: 10.1016/S0016-7037(99)00197-0
[16] 陈萍, 方念乔, 胡超涌. 有孔虫壳体Sr/Ca对冰川旋回中海平面变化的响应[J]. 安徽理工大学学报: 自然科学版, 2008, 28(4): 8-11 [CHEN Ping, FANG Nianqiao, HU Chaoyong. Sr/Ca Ratio of foraminifera shell response to the change of sea level with glacial-interglacial cycles[J]. Journal of Anhui University of Science and Technology: Natural Science, 2008, 28(4): 8-11.
CHEN Ping, FANG Nianqiao, HU Chaoyong . Sr/Ca Ratio of foraminifera shell response to the change of sea level with glacial-interglacial cycles[J]. Journal of Anhui University of Science and Technology: Natural Science,2008 ,28 (4 ):8 -11 .[17] Cléroux C, Cortijo E, Anand P, et al. Mg/Ca and Sr/Ca ratios in planktonic foraminifera: Proxies for upper water column temperature reconstruction[J]. Paleoceanography, 2008, 23(3): PA3214.
[18] Hilde T W C, Lee C S. Origin and evolution of the west Philippine basin: a new interpretation[J]. Tectonophysics, 1984, 102(1-4): 85-104. doi: 10.1016/0040-1951(84)90009-X
[19] Jia Q, Li T G, Xiong Z F, et al. Hydrological variability in the western tropical Pacific over the past 700 kyr and its linkage to Northern Hemisphere climatic change[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 493: 44-54. doi: 10.1016/j.palaeo.2017.12.039
[20] Locarnini R A, Mishonov A V, Antonov J I, et al. World Ocean Atlas 2013, volume 1: temperature[M]//Levitus S, Technical A M. NOAA Atlas NESDIS 73. NOAA, 2013.
[21] Barker S, Greaves M, Elderfield H. A study of cleaning procedures used for foraminiferal Mg/Ca paleothermometry[J]. Geochemistry, Geophysics, Geosystems, 2003, 4(9): 8407.
[22] Lisiecki L E, Raymo M E. A Pliocene‐Pleistocene stack of 57 globally distributed benthic δ18O records[J]. Paleoceanography, 2005, 20(1): PA1003.
[23] Thompson P R, Bé A W H, Duplessy J C, et al. Disappearance of pink-pigmented Globigerinoides ruber at 120, 000-yr Bp in the Indian and Pacific Oceans[J]. Nature, 1979, 280(5723): 554-558. doi: 10.1038/280554a0
[24] Brown S J, Elderfield H. Variations in Mg/Ca and Sr/Ca ratios of planktonic foraminifera caused by postdepositional dissolution: evidence of shallow Mg-dependent dissolution[J]. Paleoceanography, 1996, 11(5): 543-551. doi: 10.1029/96PA01491
[25] Elderfield H, Cooper M, Ganssen G. Sr/Ca in multiple species of planktonic foraminifera: Implications for reconstructions of seawater Sr/Ca[J]. Geochemistry, Geophysics, Geosystems, 2000, 1(11): 1017.
[26] Kısakürek B, Eisenhauer A, Böhm F, et al. Controls on shell Mg/Ca and Sr/Ca in cultured planktonic foraminiferan, Globigerinoides ruber (white)[J]. Earth and Planetary Science Letters, 2008, 273(3-4): 260-269. doi: 10.1016/j.jpgl.2008.06.026
[27] Dissard D, Reichart G J, Menkes C, et al. Mg∕Ca, Sr∕Ca and stable isotopes from the planktonic foraminifera T. sacculifer: testing a multi-proxy approach for inferring paleotemperature and paleosalinity[J]. Biogeosciences, 2021, 18(2): 423-439. doi: 10.5194/bg-18-423-2021
[28] Martin P A, Lea D W, Mashiotta T A, et al. Variation of foraminiferal Sr/Ca over Quaternary glacial-interglacial cycles: Evidence for changes in mean ocean Sr/Ca?[J]. Geochemistry, Geophysics, Geosystems, 2000, 1(12): 1004.
[29] Stoll H M, Schrag D P, Clemens S C. Are seawater Sr/Ca variations preserved in Quaternary foraminifera? [J], Geochimica et Cosmochimica Acta, 1999, 63(21): 3535-3547.
[30] Kiefer T, Kienast M. Patterns of deglacial warming in the Pacific Ocean: a review with emphasis on the time interval of Heinrich event 1[J]. Quaternary Science Reviews, 2005, 24(7-9): 1063-1081. doi: 10.1016/j.quascirev.2004.02.021
[31] Lea D W, Martin P A, Pak D K, et al. Reconstructing a 350 ky history of sea level using planktonic Mg/Ca and oxygen isotope records from a Cocos Ridge core[J]. Quaternary Sci Rev, 2002, 21(1-3): 283-293. doi: 10.1016/S0277-3791(01)00081-6
-




图(5)
表(1)
计量
- 文章访问数: 1758
- PDF下载数: 74
- 施引文献: 0